Neural Development
We are interested in understanding how cell fate is determined during neuronal development. To this end, we have studied the Notch Signalling pathway, which has been shown to detemine the fate of virtually every cell type in many complex organisms. Specifically, we have characterized a novel component of this pathway, neuralized, which encodes an E3 ubuquitin ligase that is requred for endocytosis of the Notch ligands Delta and Serrate. Current efforts are aimed at understanding how Neuralized is regulated during development and elucidating novel roles for Neuralized in cell polarity and germ line stem cell maintenance.
Key References:
Liu, S., Bonner, J.M., Chanet, S., Commisso, C., Skwarek, L.C., Schweisguth, F., Boulianne, G.L. (2012). Functional analysis of the NHR2 domain indicates that oligomerization of neuralized regulates ubiquitination and endocytosis of delta during notch signalling. Mol Cell Biol 32(24), 4933-4945.
Skwarek, L.C., and Boulianne, G.L. (2009). Great expectations for PIP: phosphoinositides as regulators of signaling during development and disease. Dev Cell 16, 12-20.
Commisso, C. & Boulianne, G.L. (2007). The NHR1 domain of Neuralized binds Delta and mediates Delta trafficking and Notch signalling. Mol. Biol Cell 18(1), 1-13.
Skwarek, L.C., Garroni, M.K., Commisso, C. & Boulianne, G.L. (2007). Neuralized contains a phosphoinositide-binding motif required downstream of ubiquitination for Delta endocytosis and Notch signalling. Dev Cell 13, 783-795.
Yeh, E., Dermer, M., Commisso, C., Zhou, L., McGlade, J. & Boulianne, G.L. (2001). Neuralized functions as an E3-ubiquitin ligase during Drosophila development Current Biology 11, 1675-1679.
For more detail see Yeh et al, 2000
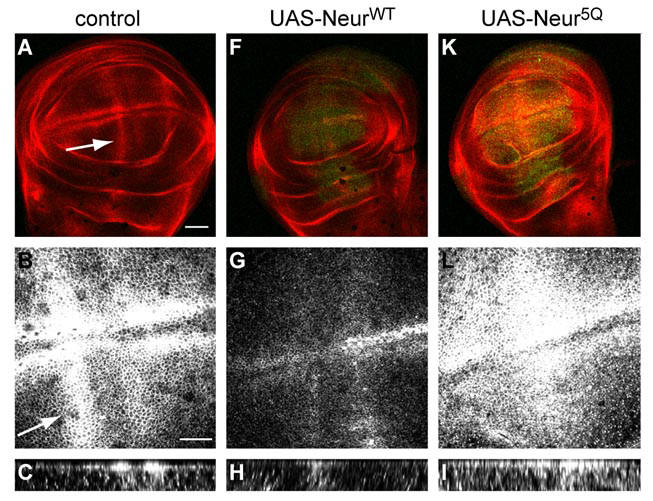
Interaction between neuralized and phospho-inositides (PIPs) is essential for endocytosis of the Notch ligand Delta. In wildtype imaginal wing discs (structures in larvae that will eventually develop into a wing), the Notch ligand Delta is expressed on the apical surface of the cells (A-C). The apical localization of Delta is clearest in the Z-stack projection in C, where strong Delta staining is visible at the top of the Z-stack. Overexpression of a wildtype neuralized construct leads to Delta internalization and a reduction in expression of Delta at the apical surface (F-H). However, overexpression of a Neuralized construct that does not bind to PIPs (Neur 5Q) fails to internalize Delta in the wing discs (K-I) suggesting that an interaction between Neuralized and PIPs is essential for Delta endocytosis, and thus Notch signalling.
Figure courtesy of Lara Skwarek. For more detail, see Skwarek et al, 2007.
Research
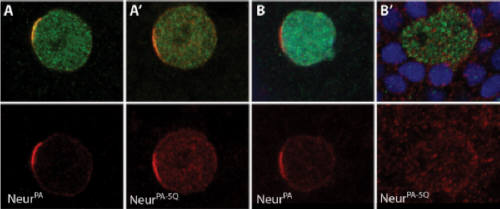
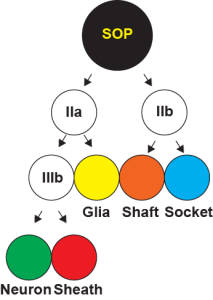
During development, a single cell can give rise to two different cells through asymmetric cell division. The bristles on a fly notum are peripheral sense organs comprised of five different cell types (hair, socket, sheath, nerve and glia). All five of these cells are derived from a single precursor cell known as the sense organ precursor (SOP). Notch signalling has been shown to be important at several steps in the development of the bristle sense organ. Shown to the left is a schematic of the cell divisions that lead from a single SOP to a fully developed sense organ. Neuralized is a key regulator of Notch signalling, and is normally localized to the plasma membrane. During SOP cell division, however, Neuralized becomes localized to the anterior side of the SOP just prior to cell division. Following division of the SOP, Neuralized is observed in the pIIa daughter cell, but not the pIIb cell. Panel A, on the right shows wild type Neuralized localizing to the anterior crescent of the SOP just prior to cell division. The SOP marker senseless is shown in green, and Neuralized is in red. Previous work in our lab has shown that a Phospho-inositide (PIP) binding region is important for cell membrane localization (Skwarek et al, 2007). Mutation of the PIP binding region in a wild-type background (A') does not affect the asymmetric localization of Neuralized in the SOP. However, in a Neuralized mutant background (B and B'), wild-type Neuralized localized asymmetrically, but a PIP binding mutant Neuralized (B') does not. These results show that the PIP binding Neuralized mutant can only localize correctly in the presence of wild-type Neuralized, suggesting that Neuralized oligomerizes.
For more information, see
Liu et al 2012
.